
Мы попытаемся дать краткое описание современного состояния этих исследований и попутно проследить, как исторически возникли идеи, послужившие основой для современного прогресса в этой области науки.
В основном здесь будет идти речь об одном из разделов молекулярной биологии, известном как молекулярная биофизика, которая имеет дело главным образом с физикой биомолекул и их ролью в объяснении химических процессов в биологических системах.
Биохимия как самостоятельная отрасль биологии возникла в начале этого столетия. Интересно заметить, что Индийский институт науки, членом которого является автор, имел биохимический отдел уже в 1920 г., когда во многих лабораториях Европы и США биохимию еще не считали самостоятельной отраслью биологии. Биохимики стали изучать химические реакции, происходящие в живых системах, и очень скоро обнаружили, что они имеют сходство с реакциями, осуществляемыми в пробирках, хотя между ними есть и ряд важных различий, обусловленных главным образом большей сложностью реакций в живых организмах. Например, для живых систем возможны реакции, которые в условиях .пробирки не могут проходить без значительного повышения температуры. Однако химикам с давних времен известны способы, позволяющие облегчить протекание реакций. Для этого используются различные катализаторы. Специфические катализаторы, управляющие реакциями в живых системах, называются ферментами. Механизм действия ферментов исследовался весьма обстоятельно. Очень скоро стало известно, что ферменты, по существу, построены из тех же самых химических материалов, из которых формируются «строительные блоки» живых организмов в животном мире, а именно — из белков. В то же время для них характерно многообразие, которое делает возможным для однотипных в химическом отношении соединений выполнение всего комплекса процессов, связанных с жизнедеятельностью. В самом деле, белки, например, образуют структурную основу живого организма, составляя его самую существенную часть, которая подвержена метаболизму, т. е. обмену веществ, и в то же время они являются соединениями, ответственными за поддержание жизненных процессов. Кроме метаболизма и роста существует еще одна важная особенность, характерная для живых систем, — способность к репродукции, т. е. самовоспроизведению.
Как известно, наследственные особенности организма связаны с ядром клетки, с содержащимися в нем хромосомами. Хромосомы состоят из соединений двух типов: белков и нуклеиновых кислот. Таким образом, белки являются важными компонентами наследственного ашвдрата клетки.
Как проявляется специфическая биологическая активность молекул, почему они ведут себя так, как это необходимо для протекания определенных процессов? Например, как ферменты ускоряют реакции? Каким образом нуклеиновые кислоты могут хранить информацию, которая преобразуется в форму, определяющую течение жизненных процессов в данной системе, и которая в конечном счете приводит к возникновению дупли-катной копии самого организма? Оказывается, изучение этих явлений тесно связано с рядом других интересных биологических проблем, таких, как возникновение болезней и резистентности по отношению к ним, т.е. с проблемами иммунологии, с исследованиями механизма действия ядов и т. д.

Рис. 1. Механизм действия антител, основанный на представлении о комплементарной форме патогенного тела, и ловушки антитела, которая его захватывает. Три разные молекулы, имеющие одинаковые по форме участки структуры, могут захватываться таким антителом
Все это при детальном рассмотрении может быть сведено к одной из проблем молекулярной физики — к проблеме соответствия одних молекул другим. Молекулярное соответствие давно связывалось с широким кругом различных явлений и в первую очередь с явлением иммунитета — свойством антител подавлять активность особых патогенов. Основная идея состояла в том, что антитело будет иметь форму, комплементарную активному центру патогена (рис. 1), и, закрывая этот активный центр, будет лишать его патогенных свойств. Но это особое свойство можно рассматривать не только как свойство биологической системы — можно также свести проблему на молекулярный уровень. В самом деле, две разные комплементарные по своей форме молекулы могут подходить друг к другу наподобие ключа и замка. Представление о таком механизме узнавания может быть использовано для объяснения очень широкого круга явлений; и мы попытаемся остановиться на принципиальных особенностях этого механизма, если говорить главным образом о биомолекулах. Мы приведем несколько примеров того, как использование этих представлений позволило объяснить сущность некоторых важных биологических явлений.
Основные особеннсти конформации биополимеров
Существует одна физическая концепция, которая является общей для всех упоминавшихся выше аспектов биологической активности. Она связана с общей формой молекул, с законами, управляющими ее вариациями.
В этой связи необходимо всегда помнить, что большинство биологически важных соединений является полимерами, т. е. химическими соединениями, в которых особые структурные единицы, присоединяясь одна к другой, образуют цепочку. Те цепные молекулы, которые имеют важнейшее биологическое значение, являются «линейными» — это означает, что при их образовании структурные единицы присоединяются одна к другой последовательно. Эти единицы для белков и нуклеиновых кислот представлены на рис. 2.
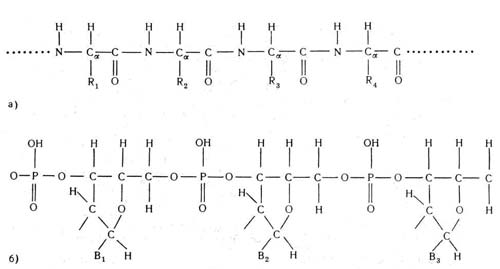
Рис. 2, Повторяющиеся группы в цепях: а — белков; б — нуклеиновых кислот. Боковые группы R1,R2 и R3 могут быть различными в белковых цепях, так же как основания В1 и В2 и В3 в нуклеиновых кислотах
Структурные единицы белковых цепей (рис. 2а) называют пептидными, они слегка отличаются от так называемых аминокислотных остатков (рис. 4), обычно используемых в органической химии при описании цепей белков. Для понимания процесса свертывания белковой цепи и результирующих изменений в конфигурации и форме молекулы наиболее удобно в качестве структурной единицы рассматривать именно пептидную единицу.
Цепь нуклеиновой кислоты (рис. 2, б) представлена схематически, без указания детального расположения атомов в каждой из групп. Важной особенностью структурных единиц как белков, так и нуклеиновых кислот является то, что они вовсе не обязаны быть идентичными. Так, группы R на рис. 2, а и группы В на рис. 2, б отличаются при переходе от одной структурной единицы к другой, в то время как все другие атомы остаются одинаковыми во всех единицах (см. также рис. 3).
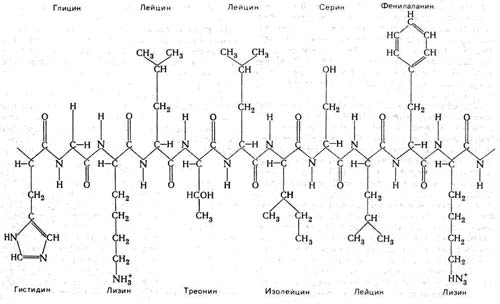
Рис. 3. Подробная схема структуры одного из участков белковой цепи миоглобина, с указанием названий различных аминокислотных остатков. Можно видеть различие структуры боковых цепочек, этим определяются химические особенности каждого белка с определенной последовательностью аминокислотных остатков
Благодаря таким вариациям структурных единиц для их достаточно большого числа можно иметь практически бесконечный набор различных цепочек, отличающихся последовательностью индивидуальных звеньев. Именно в этом и состоит причина безграничного разнообразия, характерного для живой природы.
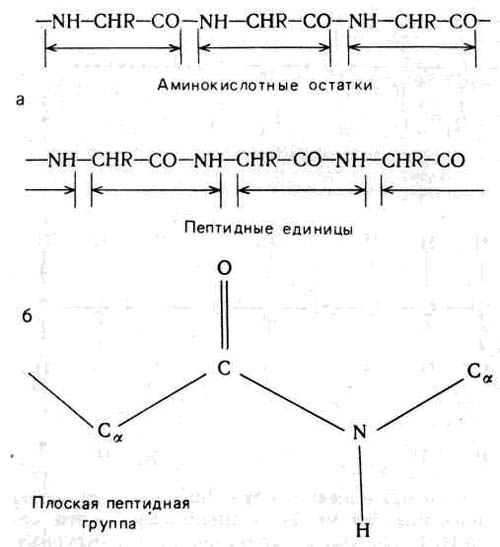
Рис. 4. Повторяющиеся структурные единицы белковой цепи (а). Вверху: аминокислотный остаток — единица, используемая для описания химических свойств белка; внизу — пептидная единица, используемая для описания стереохимии белков. Детальная схема структуры изолированной плоской пептидной группы (б)
Давайте попытаемся проследить, каким образом свертывается цепочка белка. Основные принципы этого свертывания можно понять, рассматривая возможные взаимные ориентации лишь двух структурных единиц цепочки и их изменения.
Расположение атомов остова цепи дипептидной группы, т. е. двух последовательных пептидных единиц с боковой группой, присоединенной н среднему Сжатому углерода, изображено на рис. 5.
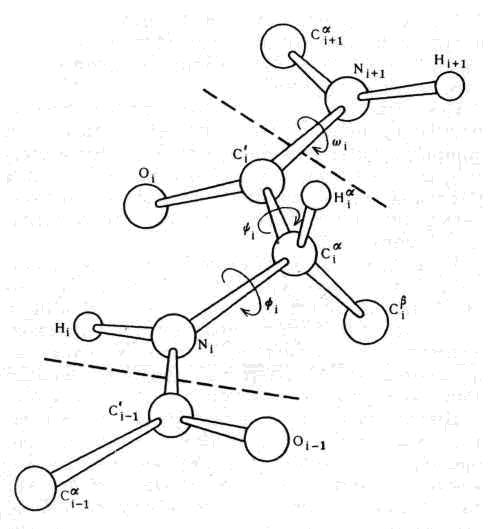
Рис. 5. Расположение атомов в двух связанных друг с другом пептидных единицах. Аминокислотный остаток обведен пунктирной линией. Хотя обе пептидные группы являются плоскими и абсолютно жестокими, их можно поворачивать вокруг связей, которыми они соединяются между собой. Два отмеченные угла поворота обычно обозначаются символами Y и Ф
Как стало известно из классических работ Лайнуса Полинга, атомы, образующие остов пептидной единицы, т. е. атомы плоской пептидной группы, все лежат в одной плоскости, что и представлено на рисунке. Концепция так называемой планариости (от английского слова plane — плоскость) пептидных групп является основным принципом структуры белков. Очевидно, следующей ступенью будет анализ того, как соединяются две пептидные группы и какие возможные взаимные пространственные ориентации они могут иметь.
Эта проблема очень подробно изучалась в нашей лаборатории в Мадрасе в 1961 — 1963 гг. Так как атомы остовов двух пептидных групп образуют жесткие планарные группировки, единственной степенью свободы при их сочленении в Сa-углеродном атоме будет поворот вокруг одинарных связей N—Сa и Сa — С’. Углы поворота, так называемые двугранный и торзионный углы, обозначаются символами Ф и Y соответственно. Легко показать, что не все комбинации Ф и Y; оказываются разрешенными, так как атомы имеют конечные размеры и начинают налезать друг на друга, если расстояние между ними оказывается меньше определенной величины, характерной для данной пары атомов. Эти лимитирующие межатомные расстояния можно определить при анализе кристаллических структур органических соединений, родственных белкам; их можно затем использовать для нахождения ограничений на возможные значения комбинации Ф и Y. Когда это было сделано и результаты были представлены в виде графика, изображенного на рис. 6, где по горизонтальной оси отложены углы Ф, а по вертикальной оси — Y мы получили так называемую «конформационную карту» пары пептидных единиц, которая содержит области разрешенных и запрещенных комбинаций Ф и Y, ограниченные линиями. Дальнейшие уточнения этих построений, на которых мы коротко остановимся позднее, позволили лучше понять, как распределяются области разрешенных конформаций для таких дипептидных единиц. Простая идея использования межатомных контактов для получения критерия разрешенных конформаций очень широко использовалась в последующих исследованиях свертывания пептидной цепи белков, а также других цепных молекул биополимеров.
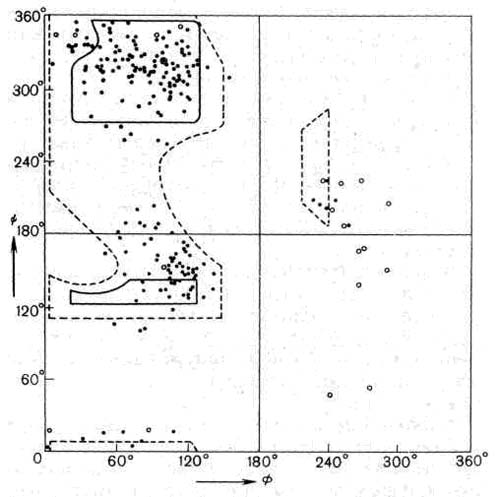
Рис. 6. Конформационная карта для пары пептидных единиц, соединяющихся Са -атомом остатка аланина, на которой изображены области разрешенных конформаций на (Y и Ф) плоскости. Полностью разрешенные области, полученные из теоретических расчетов, очерчены сплошной жирной линией, пунктиром отмечены области, соответствующие предельным значениям параметров. Точки соответствуют углам Y и Ф, известным для остатков белка химотрипсина. Светлыми кружками отмечены значения углов, характерные для остатков глицина (для которых оказывается разрешенной практически вся плоскость карты). Области локализации для 19 остатков соответствуют разрешенным областям, предсказанным теоретически (рисунок взят из статьи Бирктофта и Блоу, опубликованной в международном журнале «Молекулярная биология» в 1972 г.).
Однако продолжим наше рассмотрение и посмотрим, как такой подход позволяет понять физические принципы свертывания цепей биополимеров и связь между свертыванием и возникающими в результате химическими свойствами молекул, — теми свойствами, которые определяют их биологическую активность.
Очевидно, если можно установить разрешенные комбинации углов Ф и Y для двух соседних пептидных единиц, те же самые принципы можно использовать для каждой из пар в цепи и, таким образом, найти, какая конформация или какие изгибы возможны для длинной цепочки.
Однако оказывается, что при этом могут взаимодействовать не только соседние единицы, но и единицы, находящиеся на достаточно большом расстоянии друг от друга, что добавляет новые ограничения на возможные изгибы цепочки. Эти взаимодействия очень трудно описать математически, однако регулярность, характерная для процесса свертывания цепей биополимеров, облегчает эту задачу. Поэтому мы рассмотрим здесь те регулярные структуры, которые оказываются возможными для цепей биополимеров, и в первую очередь для белков, т. е. для полипептидов.
Спиральные структуры
Предположим, что локальный изгиб цепочки в каждом Сa -атоме, связывающем две структурные единицы, является одинаковым по всей длине полипептидной цепи. Это означает, что расположение любой из пептидных единиц по отношению к предыдущей является таким же, как расположение последующей единицы относительно рассматриваемой, поэтому такая структура в целом должна иметь определенную регулярность. Простая геометрическая теорема, которую мы здесь не будем рассматривать, показывает, что следствием регулярности локальных изгибов цепочки является образование спиральных структур, наиболее распространенных среди биополимеров.
Спираль, характерная для полипептидной цепи и обладающая высокой степенью стабильности (так называемая a-спираль), впервые предложена Полингом. Особая стабильность а-спирали является следствием образования вторичных связей, известных под названием водородных и имеющих высокую энергию стабилизации, связей между группами NH и СО остова пептидной цепи соседних витков спирали.

Рис. 7. Схема расположения атомов в одном из слоев fi-структуры. Направление следующих друг за другом полимерных цепей попеременно чередуется, они связаны водородными связями, направленными перпендикулярно длине цепочек
Свертывание цепи такого типа — весьма распространенное явление не только для фибриллярных белков, таких, как белки шерсти, волос, мышц и т. д., но и для отдельных участков молекулярной структуры биологически активных белков, таких, как гемоглобин и ферменты.
По-видимому, энергетические расчеты стабильности структуры биополимеров имели наибольший успех при предсказании точных значений Ф и Y для наиболее стабильной кон-формации спирали поли-l-аланина. Они были проведены в нашей лаборатории главным образом в работах доктора Чандрасекарана.
Другой тип регулярной структуры, так называемые b-структуры, встречающиеся, например, в волокнах шелка, растянутых волокнах шерсти и волос, был исследован различными авторами. В этом случае наблюдаемая конформация цепи в шелке и растянутой форме кератина шерсти также очень хорошо согласуется с теоретическими предсказаниями. Этот тип структуры образует основу активного центра целого ряда ферментов.
Коллаген
Третий тип спиральной структуры, возможной для белковых цепей, наблюдается в молекулах структурного белка всех соединительных тканей, известного под названием коллаген. Исследование молекулярной структуры коллагена более чем в течение 10 лет было основным направлением работы автора в лаборатории в Мадрасе. Поэтому коллаген мы (рассмотрим здесь несколько более подробно.
В 1952 г. автор возглавил вновь организованный исследовательский физический факультет в университете Мадраса и приступил к выбору направления исследований. Автор получил хорошее образование в области кристаллографии и имел достаточный опыт определения структуры веществ на молекулярном уровне, однако не имел пристрастия к изучению структуры соединений какого-то определенного типа.
Это было как раз то время, когда появились прекрасные работы Полинга и его коллег, где для разных белков предлагались различные модели структуры и рассматривались некоторые новые типы агрегации белковых молекул.
Естественно, эти работы привлекли внимание автора, поскольку они открывали ряд новых возможностей для исследований. По счастливой случайности на биохимическом факультете университета в Мадрасе оказались образцы эластоидина. Эластоидин — это одна из разновидностей коллагена, находящегося в плавниках акулы. На биохимическом факультете был определен аминокислотный состав этого вещества, и он оказался очень сходным с аминокислотным составом коллагена рыб.
Когда в лаборатории автора получили рентгенограмму этого образца, то обнаружили, что она очень похожа на рентгенограммы коллагена млекопитающих, опубликованные ранее профессором Астбери и другими исследователями.
Имея в руках свою собственную рентгенограмму, можно было провести измерения и более тщательно оценить соответствие различных структурных моделей коллагена рентгенограммам, которые для него характерны.
Было обнаружено, что хотя многие из предлагаемых структур, опубликованных в литературе, имеют особенности, согласующиеся с наблюдаемыми рентгенограммами, ни одна из них не соответствует одновременно всем данным рентгенографических и оптических исследований коллагена. Далее был отмечен один интересный момент, а именно оказалось, что для всех позвоночных остатки глицина составляли всегда приблизительно одну треть всех аминокислотных остатков коллагена. Резонно было предполагать, что такое количество остатков глицина обусловлено особыми причинами и что оно должно фиксироваться требованиями, определяемыми самой молекулярной структурой.
Хотя в ранних работах, таких, как работы Астбери, указывалось, что глицин может быть каждым третьим остатком в цепи, это требование не накладывалось самой структурой, и структура не давала объяснения, почему каждый третий остаток в цепи обязательно должен быть глицином. Автором были проанализированы различные структуры, которые можно было бы построить из спиралей с целью проверки, не получится ли что-либо подходящее для коллагена. И тогда было найдено, что одиночные спирали или пары спиралей нельзя расположить друг относительно друга, так, чтобы положение каждого третьего остатка в цепи чем-то особенным выделялось. Минимальное количество спиралей, необходимое для того, чтобы положение каждого третьего остатка отличалось от положений других остатков, оказалось равным трем. Расположение трех спиралей, полученное таким образом, изображено на рис. 8.

Рис. 8. Схема первоначальной модели коллагена, состоящей из трех спиральных цепочек, агрегированных боковыми поверхностями. Атомы Ca углерода изображены здесь кружками, а пептидные единицы — прямыми линиями, соединяющими эти атомы. Положение атомов по высоте вдоль оси спирали в ангстремах указано в скобках. Индексы при атомах углерода обозначают последовательности расположения этих атомов
Это проекция структуры остовов трех цепей на плоскость, перпендикулярную оси спирали, где кружками отмечено только положение Сa-углеродных атомов, а пептидные группы изображены в виде линий, соединяющих эти атомы. Положение Са-углеродов по высоте вдоль оси спирали в ангстремах указано в скобках.
Глядя, например, на цепь А, расположенную справа, видим, что Сa -углероды — С2 и С3 могут присоединять один водород и b-углеродный атом боковой цепи, в то время как атом Ci может присоединить только два водорода, потому что для большого р-углеродного атома здесь уже нет места. Спирали соединяются друг с другом СО—HN водородными связями, приблизительно перпендикулярными оси, что фиксирует расстояние между ними, а это расстояние исключает возможность присоединения b-углеродных атомов к атомам C1, С4 и т. д. в каждой цепочке. Другими словами, сама геометрия трехцепного комплекса требует, чтобы каждым третьим остатком в любой группе из трех пептидных единиц был бы глицин (который в качестве боковой цепи имеет лишь один атом водорода). Боковые цепи пролина и оксипролина, которых много в коллагене и которые содержат пятичленные кольца, могут располагаться при обоих атомах С2 и С3, одно из колец изображено на рис. 8.
Интересно заметить, что ориентация остова цепи при этом автоматически соответствует тому, что необходимо для присоединения подобных колец, двугранные углы, о которых говорилось выше, оказываются близкими к 100°, что точно равно величине, необходимой для такого присоединения.
Спиральная спираль
Сопоставление предложенной структуры, о которой говорилось в предыдущем разделе, с рентгенограммами показало, что она в общем соответствует эксперименту, однако это соответствие не является абсолютным. В действительности, тщательные расчеты количества структурных единиц в одном обороте
спирали, которые могут быть произведены на основе измерений рентгенограмм, дают величину, близкую к 3,3, а не 3. Таким образом, спираль оказывается, как говорят, нецелочисленной. Одна из таких нецелочисленных спиралей представлена на рис. 9а; три из них, сложенные вместе, показаны на рис. 9б. Однако на последнем рисунке можно видеть, что замечательное свойство такого комплекса, о котором говорилось выше, а именно — каждый третий Сa-углеродный атом, попадая внутрь спирального комплекса, может быть только a-углеродом глицина, оказывается здесь утерянным. Это обстоятельство принесло немало беспокойства автору и его коллеге доктору Гопинату Картха, в сотрудничестве с которым проводились эти исследования.
Идея разрешения этого кажущегося противоречия возникла лишь после длительных размышлений.
Она появилась на основе аналогий, заимствованных, как ни странно, из совершенно другой области знаний, — именно из астрономии. Хорошо известно, что Луна, вращаясь вокруг Земли, всегда остается обращенной к ней одной стороной. Причиной этого является вращение Луны вокруг собственной оси с той же скоростью, с которой она вращается вокруг Земли. Если эту идею использовать для решения проблемы коллагена, то можно получить такое расположение трех спиралей, при котором каждый третий остаток в каждой спирали (точнее, атомы 1, 4, 7) будет направлен к центру комплекса, поворачивая спирали вокруг общей оси для компенсации отклонения от 360°, возникающего вследствие того, что в каждой спирали количество остатков, приходящихся на один оборот, несколько больше, чем три.
Каждая цепь будет уже не просто спиралью, как показано на рис. 9б, она должна стать спиральной спиралью, т. е. атомы Р1 Р4 и т. д. должны находиться на поверхности цилиндра с осью, проходящей через 0, причем угол поворота от одного атома к другому должен быть постоянной величиной (рис. 9в).
Чертеж полной структуры, состоящей из трех дополнительно скрученных спиралей, расположенных соответствующим образом, представлен на рис. 9в. Здесь Сa -углеродные атомы 1, 4, 7 каждой из трех спиралей занимают эквивалентные положения внутри комплекса, поэтому необходимо, чтобы эти a-углероды принадлежали глициновым остаткам.
В то же время такая структура соответствует требованиям, накладываемым рентгеновскими дифракционными данными, а именно — ее атомы располагаются вдоль нецелочисленной спирали, имеющей приблизительно 3,3 остатка на оборот.
Можно также видеть на примере цепочки А, что b-углеродные атомы остатков пролина и оксипролина могут беспрепятственно присоединяться к a-углеродам в 2 и 3 положениях.
Исследования, о которых здесь говорится, были проведены в лаборатории автора в университете Мадраса в течение 1954—1955 гг., и разработанная таким образом модель структуры была широко признана. Обзор современных представлений о структуре коллагена был опубликован автором в книге, озаглавленной «Все о коллагене», вышедшей под его редакцией в 1966 г., а также в его лекции, посвященной памяти Джона Артура Вильсона, в американской Ассоциации химиков кожевенной промышленности. Трехспиральная модель коллагена согласуется также и с биологическими исследованиями, которые показали, что каждая цепь молекулы коллагена синтезируется независимо, и три цепи, соединяясь друг с другом, образуют жесткий трех-спиральный комплекс; он является биологически необходимым и важным компонентом коллагена.

Рис. 9. Одиночная спираль, имеющая приблизительно 3 и 1/3 остатка в одном обороте (а). Три такие спирали, агрегированные боковыми поверхностями (б); атомы а-углерода 4, 7 и т. д. все больше и больше удаляются от середины трехспирального комплекса. Три спиральные спирали, агрегированные боковыми поверхностями (в). Атомы a-углерода 1, 4, 7 и т. д. все находятся внутри трехцепного комплекса. Это основная модель трехспиральной структуры коллагена
На рис. 10 представлено фото модели остова трехспиральной структуры коллагена. Самые последние данные по анализу последовательности аминокислот в одной из трех цепей коллагена свидетельствуют, что за исключением короткого участка в начале цепи, известного под названием тело пептида, каждый третий остаток в цепочке является глицином в точном соответствии с теоретически разработанной моделью трехцепной спирали.

Рис. 10. Фотография проволочной модели трехцепной структуры коллагена. Стержни с нанесенными полосками изображают водородные связи между цепочками. Шариками представлены молекулы воды, которые участвуют в образовании таких связей (из работы Рамачандрана и Чандра-секхарана, опубликованной в журнале «Биополимеры» в 1968 г.)
Трехцепная спираль, имеющая приблизительно 1000 остатков аминокислот в каждой цепи и молекулярный вес около 300 000, является «строительным блоком» коллагеново-го волокна.
Структура глобулярных белков
Выше мы рассмотрели молекулярную кон-формацию так называемых структурных белков, т. е. тех белков, из которых формируется структурная основа живого организма, т. е. мышцы, сухожилия, кости и т. д. Белки, входящие в состав этих тканей, имеют преимущественно регулярную спиральную структуру одного из трех рассмотренных типов: a-спираль, b-структуру или трехцёпную спираль коллагена. Однако цепи, содержащие такие спиральные структуры, могут иметь различные молекулярные веса и соответственно разное число пептидных единиц. С другой стороны, так называемые глобулярные белки, участвующие в физиологических процессах живого организма, имеют вполне определенные молекулярные веса. Они обычно содержат цепочки, состоящие от 50 до 300 остатков аминокислот, и имеют молекулярные веса от 5000 до 30 000 и более.
В этих белках нет особой заметной регулярности чередования остатков аминокислот в отличие от коллагена, у которого каждый третий остаток является глицином. Поэтому их молекулярные цепи скручиваются нерегулярным образом. Однако сворачивание оказывается идентичным для всех молекул одного и того же белка в различных организмах, Другими словами говоря, хотя скручивание цепочки нерегулярно, оно не является хаотическим и следует одному и тому же закону для всех молекул данного белка, находящихся в одинаковых физиологических условиях. В то же время в свернутой цепочке можно обнаружить отдельные локальные участки с регулярной структурой, как a-спираль или b-структура. Например, в молекулах таких важных белков, как миоглобин и гемоглобин, значительное количество участков имеют форму a-спирали.
Ферменты, такие, как химотрипсин и па-паин, содержат значительное количество участков цепи в (3-конформации, причем эти участки располагаются друг относительно друга наподобие антипараллельной укладки цепочек.
Мы не будем углубляться здесь в детали этого вопроса, укажем лишь, что даже в белках, молекулы которых свернуты сложным образом и имеют компактную, почти сферическую форму, тенденция к образованию регулярных структур благодаря их высокой степени стабильности остается весьма заметной.
Вследствие характерного свертывания длинных цепочек метаболических белков, разные группы, принадлежащие различным участкам цепи, образующим активный центр молекулы, оказываются в непосредственной близости друг от друга. В своей совокупности они принимают участие в процессах, связанных с биологической активностью белка.
Поэтому очевидно, что биологические функции данного белка в системе определяются в немалой степени его молекулярной структурой. Для того чтобы полностью понять различные факторы, определяющие биологическую активность белков, необходимо в первую очередь проанализировать молекулярные силы, которые приводят к возникновению активной конформации, принимаемой белковой молекулой. Ряд лабораторий в различных странах мира заняты сейчас этой проблемой, причем многие исследования требуют применения мощных вычислительных машин.
Структура нуклеиновых кислот и молекулярная генетика
Интерес к молекулярной структуре нуклеиновых кислот необычайно возрос после того, как.в 1952 г. Уотсоном и Криком была постулирована хорошо известная спиральная структура ДНК (дезоксирибонуклеиновой кислоты). Она настолько хорошо всем знакома, что мы не приводим здесь ее изображения. Интересно заметить, что структура образована двумя спиралями, которые закручены вокруг общей оси, и стабильность такой двойной спирали возрастает вследствие возникновения водородных связей между соответствующими основаниями разных спиралей, расположенными на одной высоте друг против друга. Связанные таким образом основания образуют систему комплементарных пар (следуя принятым обозначениям, так называемые AT- и GC-пары). Благодаря тому что А связывается только с Т, a G с С, становится понятным механизм репродукции ДНК, которая является веществом наследственности во всяком живом организме. Две цепи раскручиваются и расходятся, после чего каждая из них способна служить матрицей для синтеза комплементарной цепи. Поэтому две образовавшиеся молекулы ДНК являются репликами исходной молекулы. Все это оказывается возможным благодаря особенностям образования вторичных валентных — именно водородных связей.
Кроме дезоксирибонуклеиновой, существует также рибонуклеиновая кислота (РНК), которая принимает участие в метаболизме и участвует в синтезе белков. Если ДНК является веществом наследственности и последовательность оснований в ней определяет наследуемые особенности организма, то истинный механизм, с помощью которого информация, закодированная в такой последовательности, трансформируется в активность клетки, связан с функцией РНК, известной как информационная РНК (m-РНК), которая в первую очередь синтезируется с помощью соответствующего процесса копирования ДНК. Существует огромное количество различных m-РНК, соответствующих всему многообразию белков и ферментов в живых организмах. Каждый белок синтезируется на соответствующей ему информационной РНК также с помощью процесса определенной репликации, однако в данном случае приходится иметь дело с перекодировкой последовательности оснований в последовательность аминокислот. Было доказано, что кодом для каждой аминокислоты является определенная последовательность трех нуклеотидных единиц. Мы не описываем здесь подробно генетический код, так как этот вопрос широко был освещен на страницах печати и хорошо известен.
Однако если необходимо понять основы механизма синтеза РНК и синтеза белка на РНК, следует вновь обратиться к молекулярным силам, участвующим в этих процессах и обеспечивающим те конформации, которые принимают цепи. Первый анализ конформации нуклеиновых кислот для оценки двугранных углов был проведен в лаборатории автора главным образом в сотрудничестве с доктором Сасисекхараном и другими коллегами. Мы рассмотрели нуклеотидную единицу, изображенную на рис. 11, и возможные двугранные углы, образующиеся в ней.

Рис. 11. Возможные двугранные углы в нуклеотиде
С помощью рентгеноструктурного анализа было исследовано большое число нуклеотидов и ди-нуклеотидов, и теперь нам хорошо известны области значений этих углов, которые отвечают стабильной структуре нуклеиновых кислот. Интересно заметить, что теоретические выводы здесь очень хорошо согласуются с экспериментальными данными. Еще более интересным является тот факт, что тщательный анализ изолированной нуклеотидной единицы, диапазона разрешенных для ее конформации значений двугранных углов помогает понять, почему возникают определенные спиральные структуры в полинуклео-тидах и нуклеиновых кислотах.
Структура полисахаров
В то время как белки образуют главную структурную основу всех тканей животных, полисахара являются доминирующими веществами в растительном царстве. Точно так же, как белки и нуклеиновые кислоты, поли-сахара образуются при соединении друг с другом большого числа однотипных остатков (рис. 12). Однако в этом случае остатки построены из молекул Сахаров. Хорошо известный сахар — глюкоза составляет главную часть полисахаров, представляющих очень большой интерес, именно — целлюлозы, образующей основу структуры большинства растений, и крахмала, который входит в состав важных пищевых продуктов. Сахарные кольца, которые встречаются в этих полисахарах, представлены на рис. 13. Две изображенные модификации глюкозы — а-глюкоза и (3-глю-коза — содержатся соответственно в крахмале и целлюлозе.

Рис. 12. Повторяющиеся группы в цепях полисахаров

Рис. 13. Молекулярная структура a-глюкозы и b-глюкозы. Направление связи C1O1 и С4О4 приблизительно перпендикулярно в первом случае и приблизительно параллельно во втором
Схема связей, появляющихся при соединении остатков глюкозы, для обоих их типов показана на рис. 14. Можно видеть, что в случае b-глюкозы цепь имеет удлиненную, почти вытянутую конформацию. Эта конфор-мация встречается, например, в волокнах хлопка (который является одним из лучших примеров целлюлозы), а также в древесине. Высокая степень жесткости растянутой цепочки обеспечивает основной запас прочности целлюлозных волокон. Несколько волокон могут агрегировать боковыми поверхностями, вследствие чего они становятся нерастворимыми в воде. С другой стороны, при наращивании цепочки а-глюкозы, по способу соединения двух остатков, изображенному на рис. 14 а, молекулярная цепь начинает скручиваться и возникают взаимодействия между вновь образованными сегментами и сегментами в начале цепочки. Вследствие этого амилоза — полимер a-глюкозы — имеет в высшей степени свернутую структуру, что хорошо установлено экспериментальными исследованиями растворов крахмала. В действительности такие цепочки могут также циклизоваться, образуя шести- и девятичленные кольца циклической молекулы амилозы. Эти соединения также хорошо известны.

Рис. 14. Два соседних остатка a-глюкозы, ориентированных относительно друг друга в соответствии с допустимыми значениями углов вращения Ф и Y. В этом полимере цепочка может закручиваться, меняя направление цепи на противоположное и образуя таким образом растворимый крахмал (а). В случае b-глюкозы при допустимых значениях Ф и Y цепочка оказывается почти вытянутой и негибкой (б). Такие цепи агрегируют друг с другом боковыми поверхностями, образуя прочные волокна, такие, как, например, волокна древесины и других растений
Разница в соединении остатков а и b-глюкозы определяет различие физических и химических свойств важнейших веществ, содержащих глюкозу и присутствующих в растениях. Природа связей между соседними структурными единицами и образование изгибов цепей или спиральных структур в цепях полисахаров (полимеров Сахаров разных типов) исследовались в течение последних десяти лет в прежней лаборатории автора в Мадрасе, главным образом доктором В. С. Р. Рао и его коллегами. Как уже указывалось, эти теоретические исследования четко показывают, что можно ожидать или жесткой компактной структуры, подобной той, которая реализуется для целлюлозы в древесине или стеблях растений, или очень гибкой, с образованием растворимого вещества, которая характерна для амилозы или крахмала, легко перевариваемых в организме человека в противоположность целлюлозе.
В то же время пищеварительная система таких животных, как коровы, лошади и другие травоядные, содержит специальный фермент — целлюлазу, который переваривает целлюлозу, превращая ее сначала в глюкозу.
Теория также предсказывает, что в соответствующих условиях амилоза может образовать спираль, но эта спираль должйа иметь большой диаметр и малую высоту витка с большой пустой полостью в центре. Крупные атомы, такие, как атом иода, могут поместиться в эту центральную полость. Это является причиной характерной реакции крахмала на иод с образованием голубого комплекса.
Общие принципы биологической активности
Все рассмотренные примеры подтверждают нашу основную концепцию, которая говорит о том, что специфичность активности молекул биологических систем определяется их физико-химическими свойствами.
Мы видим, что биомолекулы могут свертываться различным образом, однако очень часто реализуется один специфический тип этого свертывания, соответствующий минимуму энергии. Конформация при этом минимуме обычно оказывается очень стабильной, поэтому объяснение биохимических аспектов активности этих молекул следует искать в пространственном распределении активных групп. Очень часто биологически активная , молекула, например молекула фе|рмента, не только принимает стабильную конформа-цию, когда сближаются активные группы, но приобретает также свойство притягивать молекулы субстрата, на который действует фермент. Благодаря сближению соответствующих групп фермент может «зажать субстрат в клещи».
Прекрасным примером, иллюстрирующим это явление, оказались рентгено-структурные исследования конформации фермента карбоксипептидазы, проведенные профессором Липекомбом и его коллегами в Гарвардском университете. Интересны конформационные изменения, которые происходят, когда этот фермент реагирует с карбоксильным концом белковой цепочки, отщепляя одну пептидную единицу с конца.
Когда фермент связывает пептид, вследствие возникающих взаимодействий в конформации его молекулы происходит ряд изменений. Например, бензольное кольцо боковой цепи одного из тирозиновых остатков смещается из-за поворота вокруг Сa —Сb связи примерно на 120°, при этом требуемая для активности группа в тирозиновом остатке уже принимает необходимое положение в молекуле. В действительности в нормальном состоянии фермент имеет тирозиновую боковую цепочку в конформации, соответствующей ее максимальной стабильности, как это было показано в нашей лаборатории при исследованиях структуры аминокислот и пептидов.
Однако при взаимодействии с субстратом боковая тщюзиновая цепочка принимает необычную ориентацию. В то же время это и есть та самая конформация, при которой взаимодействие с субстратом оказывается большим, т. е. она наиболее благоприятна для такого взаимодействия. Итак, различные химические стадии действия этого фермента становятся понятными, если анализировать силы молекулярных взаимодействий.
Мы стремились показать, что жизнь в своей основе не отличается от процессов, происходящих в небиологических системах, так как во всех случаях определяющими являются химические взаимодействия.
Однако системы, входящие в состав живого организма, настолько высокоорганизованы, что могут принимать участие в необычайно сложных химических реакциях, которые специфичны для этих систем и характеризуются удивительным разнообразием.
К сожалению, мы не имеем возможности углубляться в такие вопросы, как проблема обратной связи в биологических системах, которая играет доминирующую роль в контроле и комбинациях различных реакций.
Автор, однако, надеется, что аргументы, приведенные в этой статье, достаточно убедительно показывают, что нет никакой необходимости привлекать новые гипотезы, кроме тех, которые обычно используются в физических и химических лабораториях, чтобы понять причины сложности и многообразия процессов, связанных с жизнедеятельностью.
Гопаласамудрам Рамачандран
индийский биофизик.
Наука и человечество. 1975. Сборник - М., «Знание», 1974. С. 197–211