
Основные особеннсти конформации биополимеров
Существует одна физическая концепция, которая является общей для всех упоминавшихся выше аспектов биологической активности. Она связана с общей формой молекул, с законами, управляющими ее вариациями.
В этой связи необходимо всегда помнить, что большинство биологически важных соединений является полимерами, т. е. химическими соединениями, в которых особые структурные единицы, присоединяясь одна к другой, образуют цепочку. Те цепные молекулы, которые имеют важнейшее биологическое значение, являются «линейными» — это означает, что при их образовании структурные единицы присоединяются одна к другой последовательно. Эти единицы для белков и нуклеиновых кислот представлены на рис. 2.
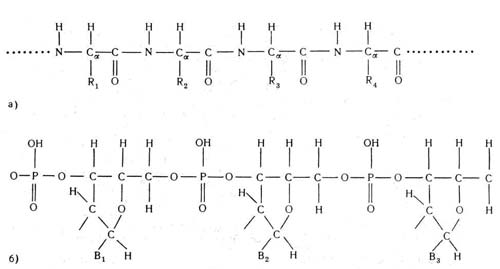
Рис. 2, Повторяющиеся группы в цепях: а — белков; б — нуклеиновых кислот. Боковые группы R1,R2 и R3 могут быть различными в белковых цепях, так же как основания В1 и В2 и В3 в нуклеиновых кислотах
Структурные единицы белковых цепей (рис. 2а) называют пептидными, они слегка отличаются от так называемых аминокислотных остатков (рис. 4), обычно используемых в органической химии при описании цепей белков. Для понимания процесса свертывания белковой цепи и результирующих изменений в конфигурации и форме молекулы наиболее удобно в качестве структурной единицы рассматривать именно пептидную единицу.
Цепь нуклеиновой кислоты (рис. 2, б) представлена схематически, без указания детального расположения атомов в каждой из групп. Важной особенностью структурных единиц как белков, так и нуклеиновых кислот является то, что они вовсе не обязаны быть идентичными. Так, группы R на рис. 2, а и группы В на рис. 2, б отличаются при переходе от одной структурной единицы к другой, в то время как все другие атомы остаются одинаковыми во всех единицах (см. также рис. 3).
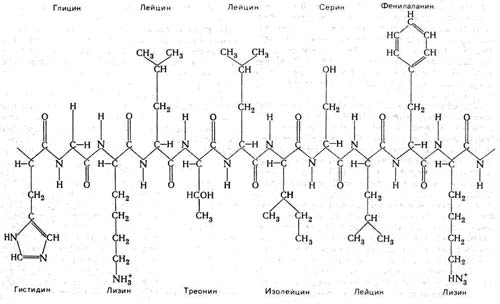
Рис. 3. Подробная схема структуры одного из участков белковой цепи миоглобина, с указанием названий различных аминокислотных остатков. Можно видеть различие структуры боковых цепочек, этим определяются химические особенности каждого белка с определенной последовательностью аминокислотных остатков
Благодаря таким вариациям структурных единиц для их достаточно большого числа можно иметь практически бесконечный набор различных цепочек, отличающихся последовательностью индивидуальных звеньев. Именно в этом и состоит причина безграничного разнообразия, характерного для живой природы.
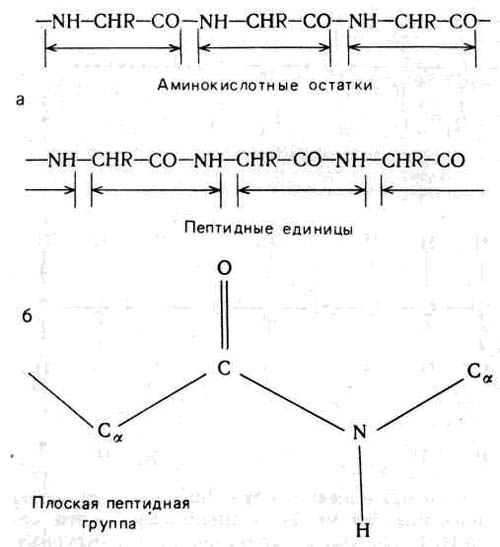
Рис. 4. Повторяющиеся структурные единицы белковой цепи (а). Вверху: аминокислотный остаток — единица, используемая для описания химических свойств белка; внизу — пептидная единица, используемая для описания стереохимии белков. Детальная схема структуры изолированной плоской пептидной группы (б)
Давайте попытаемся проследить, каким образом свертывается цепочка белка. Основные принципы этого свертывания можно понять, рассматривая возможные взаимные ориентации лишь двух структурных единиц цепочки и их изменения.
Расположение атомов остова цепи дипептидной группы, т. е. двух последовательных пептидных единиц с боковой группой, присоединенной н среднему Сжатому углерода, изображено на рис. 5.
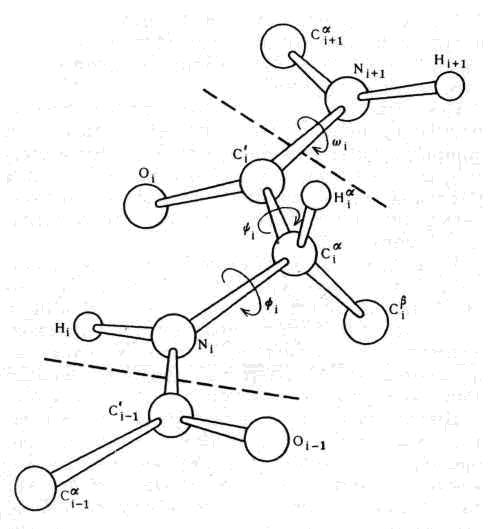
Рис. 5. Расположение атомов в двух связанных друг с другом пептидных единицах. Аминокислотный остаток обведен пунктирной линией. Хотя обе пептидные группы являются плоскими и абсолютно жестокими, их можно поворачивать вокруг связей, которыми они соединяются между собой. Два отмеченные угла поворота обычно обозначаются символами Y и Ф
Как стало известно из классических работ Лайнуса Полинга, атомы, образующие остов пептидной единицы, т. е. атомы плоской пептидной группы, все лежат в одной плоскости, что и представлено на рисунке. Концепция так называемой планариости (от английского слова plane — плоскость) пептидных групп является основным принципом структуры белков. Очевидно, следующей ступенью будет анализ того, как соединяются две пептидные группы и какие возможные взаимные пространственные ориентации они могут иметь.
Эта проблема очень подробно изучалась в нашей лаборатории в Мадрасе в 1961 — 1963 гг. Так как атомы остовов двух пептидных групп образуют жесткие планарные группировки, единственной степенью свободы при их сочленении в Сa-углеродном атоме будет поворот вокруг одинарных связей N—Сa и Сa — С’. Углы поворота, так называемые двугранный и торзионный углы, обозначаются символами Ф и Y соответственно. Легко показать, что не все комбинации Ф и Y; оказываются разрешенными, так как атомы имеют конечные размеры и начинают налезать друг на друга, если расстояние между ними оказывается меньше определенной величины, характерной для данной пары атомов. Эти лимитирующие межатомные расстояния можно определить при анализе кристаллических структур органических соединений, родственных белкам; их можно затем использовать для нахождения ограничений на возможные значения комбинации Ф и Y. Когда это было сделано и результаты были представлены в виде графика, изображенного на рис. 6, где по горизонтальной оси отложены углы Ф, а по вертикальной оси — Y мы получили так называемую «конформационную карту» пары пептидных единиц, которая содержит области разрешенных и запрещенных комбинаций Ф и Y, ограниченные линиями. Дальнейшие уточнения этих построений, на которых мы коротко остановимся позднее, позволили лучше понять, как распределяются области разрешенных конформаций для таких дипептидных единиц. Простая идея использования межатомных контактов для получения критерия разрешенных конформаций очень широко использовалась в последующих исследованиях свертывания пептидной цепи белков, а также других цепных молекул биополимеров.
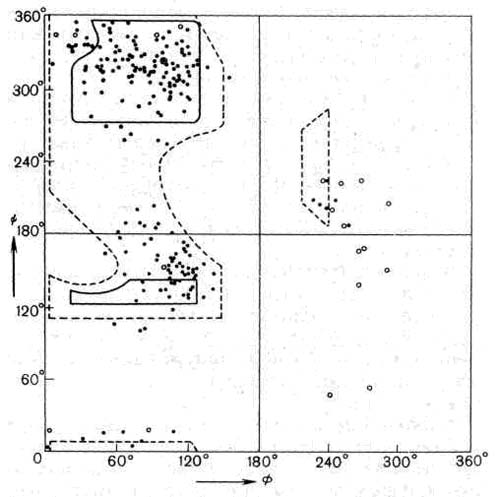
Рис. 6. Конформационная карта для пары пептидных единиц, соединяющихся Са -атомом остатка аланина, на которой изображены области разрешенных конформаций на (Y и Ф) плоскости. Полностью разрешенные области, полученные из теоретических расчетов, очерчены сплошной жирной линией, пунктиром отмечены области, соответствующие предельным значениям параметров. Точки соответствуют углам Y и Ф, известным для остатков белка химотрипсина. Светлыми кружками отмечены значения углов, характерные для остатков глицина (для которых оказывается разрешенной практически вся плоскость карты). Области локализации для 19 остатков соответствуют разрешенным областям, предсказанным теоретически (рисунок взят из статьи Бирктофта и Блоу, опубликованной в международном журнале «Молекулярная биология» в 1972 г.).
Однако продолжим наше рассмотрение и посмотрим, как такой подход позволяет понять физические принципы свертывания цепей биополимеров и связь между свертыванием и возникающими в результате химическими свойствами молекул, — теми свойствами, которые определяют их биологическую активность.
Очевидно, если можно установить разрешенные комбинации углов Ф и Y для двух соседних пептидных единиц, те же самые принципы можно использовать для каждой из пар в цепи и, таким образом, найти, какая конформация или какие изгибы возможны для длинной цепочки.
Однако оказывается, что при этом могут взаимодействовать не только соседние единицы, но и единицы, находящиеся на достаточно большом расстоянии друг от друга, что добавляет новые ограничения на возможные изгибы цепочки. Эти взаимодействия очень трудно описать математически, однако регулярность, характерная для процесса свертывания цепей биополимеров, облегчает эту задачу. Поэтому мы рассмотрим здесь те регулярные структуры, которые оказываются возможными для цепей биополимеров, и в первую очередь для белков, т. е. для полипептидов.